

Регулярная выбраковка суточных цыплят-самцов представляет собой серьезную этическую проблему в птицеводческой отрасли. Эта практика запрещена в Германии, и аналогичный запрет рассматривается Европейским союзом. Ежегодно в ЕС уничтожаются сотни миллионов суточных цыплят-самцов, а во всем мире — несколько миллиардов. Разработаны различные методы определения пола цыплят до вылупления; однако большинство из них инвазивны и позволяют определить пол относительно поздно, потенциально после начала болевого восприятия у эмбрионов. Существующие подходы включают анализ методом полимеразной цепной реакции, спектроскопию, анализ летучих органических соединений, морфологический анализ и машинное зрение.
Предыдущие исследования показали, что машинное зрение может достигать точности до 89,25% за счет анализа структуры кровеносных сосудов на ранних стадиях инкубации. Несмотря на этот потенциал, исследования остаются ограниченными, особенно в отношении различных пород кур и временного развития эмбрионов. В этом исследовании мы изучаем влияние как породных различий, так и временной информации на определение пола на ранних стадиях. Изображения были собраны на 4-й, 5-й и 6-й дни инкубации из 208 куриных яиц.
Для анализа пространственных и временных характеристик были оценены сверточная нейронная сеть (CNN) и гибридная модель сверточной нейронной сети – рекуррентной нейронной сети (CNN–RNN). Результаты показывают, что модель CNN достигла точности до 71,43%, в то время как гибридная модель CNN–RNN достигла 67,85%. Эти результаты указывают на то, что включение временной информации не улучшило производительность по сравнению с базовой моделью CNN. Однако из-за ограниченного размера и качества набора данных окончательные выводы сделать нельзя.
1. Введение
Регулярная выбраковка суточных цыплят-самцов представляет собой серьезную этическую проблему в современном птицеводстве, затрагивающую миллионы животных ежегодно [1]. Это послужило стимулом для разработки неинвазивных методов определения пола на ранней стадии, которые могут применяться до вылупления.
Было предложено несколько подходов, включая спектроскопические методы, молекулярный анализ и методы, основанные на гормонах, многие из которых обеспечивают высокую точность, но являются инвазивными, дорогостоящими или ограничены более поздними стадиями инкубации [2, 3, 4]. В последнее время машинное зрение стало многообещающей неинвазивной альтернативой, способной определять пол на ранних стадиях на основе рисунка кровеносных сосудов эмбриона [5, 6, 7]. Кровеносные сосуды эмбриона становятся четко видны примерно на 4–6 день инкубации, что делает эту стадию подходящей для анализа изображений.
Предыдущие исследования продемонстрировали обнадеживающие результаты использования моделей глубокого обучения, при этом сообщалось о точности, приближающейся к 90% в контролируемых условиях [5]. Однако эти исследования обычно основаны на однородных наборах данных, собранных в лабораторных или промышленных инкубаторных условиях, где изменчивость характеристик скорлупы яиц, условий освещения и качества изображения ограничена. В результате обобщаемость таких моделей на более реалистичные условия остается в значительной степени непроверенной в более изменчивых условиях реального мира.
Кроме того, хотя эмбриональное развитие по своей природе является временным процессом, потенциальная польза от включения временной информации в системы определения пола на основе машинного зрения получила ограниченное внимание в предыдущих работах.
В данном исследовании мы устраняем эти ограничения, изучая определение пола на ранних стадиях в более реалистичных условиях. В частности, в этой работе (i) используется гетерогенный набор данных, собранный от нескольких пород кур в реальных условиях фермы, (ii) оценивается влияние временной информации с использованием гибридного подхода на основе сверточной нейронной сети и рекуррентной нейронной сети (CNN–RNN), и (iii) оценивается обобщаемость моделей машинного зрения за пределами контролируемых экспериментальных условий.
2. Материалы и методы
Этот раздел состоит из четырех подразделов: Сбор данных, Система сбора данных, Предварительная обработка и Модели машинного обучения (МО). В совокупности они описывают получение набора данных, техническую настройку, используемую для получения изображений, этапы подготовки данных и стратегии моделирования, применяемые в данном исследовании.
2.1. Сбор данных
Исследование проводилось на ферме Экебо Фьедерфя [8] в течение восьми недель в 2024 году. Были включены три породы: Вельсумер, Крем Легбар и помесь Смоландсхёнан и Крем Легбар. Эти породы были выбраны потому, что их специфические для пола узоры оперения облегчают определение пола после вылупления, как показано на рисунке 1.

Рисунок 1. Вылупившиеся птенцы вельзумерской породы (слева) и вылупившиеся птенцы кремовой легбарской породы (справа; самец слева).
Куры породы Вельзумер, происходящие из Нидерландов, несут темно-коричневые яйца, часто с пятнистой скорлупой ( рис. 2 , справа). Они несут около 230 яиц в год, средний вес яйца составляет около 60 г. Самок обычно можно узнать по более выраженной полосе возле глаза и более четкому рисунку на голове. Куры породы Крем Легбар, выведенные в Великобритании как помесь, несут яйца с характерным голубоватым оттенком ( рис. 2 , слева). В среднем они несут около 250 яиц в год, каждое яйцо весит 55–60 г. В этой породе возможно определение пола по оперению, так как самцы имеют гораздо более светлую окраску по сравнению с самками.

Рисунок 2. Яйцо от кур породы Cream Legbar или помеси Cream Legbar/Smålandshöna (слева) и яйцо от кур породы Welsumer (справа).
Использованная в этом исследовании помесь, выведенная на ферме Экебо Фьедерфя, включает в себя шведскую породу Смоландсхёна, которая происходит от пород Леггорн и Род-Айленд Ред. Яйца этой помеси напоминают яйца породы Крем Легбар, а самцов можно отличить по тем же характеристикам оперения, что и у Крем Легбар.
Для экспериментов яйца инкубировали в инкубаторе Brinsea OvaEasy 190 Advance (Brinsea Products Ltd., Уэстон-супер-Мэр, Великобритания) [9] партиями по 25–50 яиц, каждое из которых было помечено номером партии и идентификатором яйца на тупом конце. Для сохранения естественных условий яйца не очищали перед инкубацией. Температура в инкубаторе была установлена на 37,7 °C при относительной влажности 40 %, и инкубатор вращали каждые 60 минут. Температурные параметры были выбраны на основе рекомендаций предыдущих исследований. В общей сложности для данного исследования было инкубировано восемь партий яиц.
Изображения были получены на 4, 5 и 6 дни инкубации, что на один день позже, чем в большинстве аналогичных исследований, где в качестве первой точки наблюдения обычно используется 3-й день [5, 6, 10]. Эта корректировка была необходима, поскольку в яйцах Welsumer на 3-й день ни эмбрион, ни сосудистые структуры не были четко видны. Поскольку анализ также учитывал временные аспекты с использованием рекуррентных нейронных сетей (РНН), яйца фотографировали с интервалом в 24 часа с точностью ±60 мин.
Во время получения изображений примерно треть каждой партии временно извлекали из инкубатора. Эта процедура отражает естественное поведение наседок, которые периодически покидают свои гнезда во время инкубации [11]. Чтобы минимизировать риск переохлаждения и поддерживать соответствующие температурные условия, яйца помещали на нагревательный коврик во время съемки. На рисунке 3 представлены репрезентативные изображения за 4, 5 и 6 дни, демонстрирующие прогрессирующее развитие эмбрионов и их сосудистых структур.
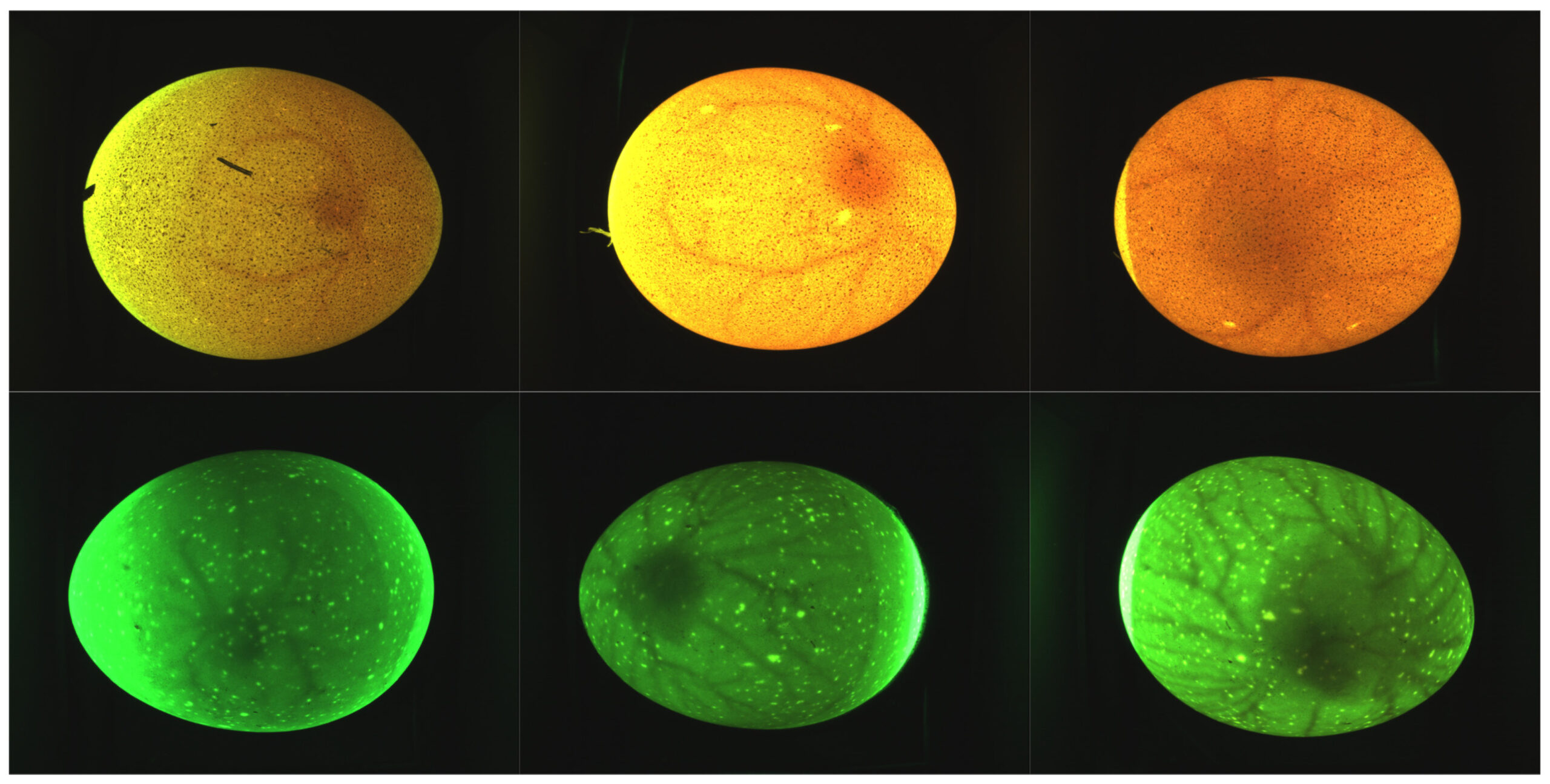
Рисунок 3. Типичные изображения яиц на разных днях инкубации: 4-й день (слева), 5-й день (посередине) и 6-й день (справа). В верхнем ряду показаны яйца овсянки сорта Вельзумер, а в нижнем — яйца овсянки сорта Крем Легбар. Все изображения были получены с использованием одного источника света — зеленого светодиода (540 нм). Различия в цветопередаче обусловлены вариациями свойств скорлупы, влияющими на светопропускание.
Качество яиц, использованных в этом исследовании, варьировалось, поскольку они были получены с небольшой фермы, а не с промышленного птицеводческого предприятия. На некоторых яйцах были заметны светлые и темные пятна на скорлупе. Приближаясь к ожидаемому дню вылупления (21-й день инкубации), примерно треть яиц из каждой партии извлекали из инкубатора для получения изображений. Для поддержания надлежащих температурных условий в течение этого процесса яйца помещали на нагревательный коврик. Для точной маркировки все яйца на 21-й день переносили в нейлоновые сетчатые мешки и маркировали их соответствующим номером партии и идентификатором яйца. Пол цыплят определяли при вылуплении (21-й день) обученным персоналом фермы Экебо Фьедерфа, а затем документировали авторы.
В общей сложности было инкубировано 271 яйцо в восьми партиях. Яйца, не показавшие признаков развития, были исключены из дальнейшего наблюдения, а 43 яйца не были определены по полу из-за неполного развития. Яйца без подтвержденной маркировки пола или без полной серии изображений за все три дня были исключены из окончательного набора данных. После этой фильтрации для анализа осталось 208 яиц с 624 изображениями (из первоначальных 790 полученных изображений). Распределение инкубированных яиц по трем породам представлено в таблице 1.
Таблица 1. Распределение инкубированных яиц по трем породам. В последних двух столбцах указано количество цыплят мужского и женского пола с подтвержденным указанием пола. В общей сложности в окончательный набор данных было включено 208 яиц, что соответствует 624 изображениям.
| Порода | Мужской | Женский | Нет настоящей метки |
|---|---|---|---|
| Велсумер | 36 | 23 | 10 |
| Кремовый Легбар | 70 | 65 | 24 |
| Скрещивание | 18 | 16 | 9 |
| Сумма | 124 | 104 | 43 |
2.2. Собранное фотооборудование
Для получения изображений во время инкубации была разработана прототипная система визуализации. Система состояла из трех основных компонентов: камеры, препарата для яиц и светодиодной матрицы для освещения (рис. 4). Камера была подключена к Raspberry Pi, который передавал изображения на компьютер для хранения в базе данных. База данных содержала информацию о породе, временной метке и партии. Кроме того, Raspberry Pi использовался для приема команд управления и пересылки полученных изображений на компьютер для последующего анализа.

Рисунок 4. Прототип устройства для получения изображений, состоящий из трех основных компонентов: камеры, держателя для яиц и светодиодной матрицы. К камере был подключен Raspberry Pi для хранения изображений и передачи данных в базу данных.
Пластиковые компоненты устройства для получения изображений были изготовлены методом 3D-печати из полимолочной кислоты (PLA). Внешние размеры устройства составляли 86 × 172 мм. Расстояние от держателя яиц до источника света составляло 56 мм, а расстояние между держателем яиц и объективом камеры — 86 мм. Держатель яиц был разработан с отверстием в форме яйца для обеспечения стабильного расположения яиц во время получения изображений. Позиционирование яиц было частично стандартизировано с помощью этого держателя, что помогло поддерживать одинаковое положение между изображениями.
Однако ориентация яиц, включая вращение и горизонтальное выравнивание, не контролировалась строго для всех образцов. Хотя изображения в рамках одной сессии получения изображений в целом были согласованными, между сессиями возникали вариации, вносящие дополнительную изменчивость в набор данных. Для получения изображений использовалась камера Raspberry Pi High Quality Camera, подключенная к Raspberry Pi Zero 2W (Raspberry Pi Ltd., Кембридж, Великобритания) вместе с массивом светодиодов.
Массив состоял из схемы с регулируемой яркостью тока и 12 зеленых светодиодов (GT PSLR31.13, OSRAM Opto Semiconductors GmbH, Регенсбург, Германия), обеспечивающих суммарный световой поток 2112 люмен и излучающих свет на длине волны 540 нм. Как отмечалось в предыдущих исследованиях, воздействие зеленого света во время инкубации может способствовать благоприятному эмбриональному развитию [12, 13, 14].
2.3. Предварительная обработка
В процессе предварительной обработки изображений необходимо было решить ряд проблем, в частности, проблему несбалансированного освещения, возникающую из-за различий в характеристиках яичной скорлупы. Предварительная обработка изображений проводилась с использованием OpenCV (версия 4.9.0) и Scikit-image (версия 0.23.2).
Основной алгоритм предварительной обработки, показанный на рисунке 5, состоял из четырех этапов:
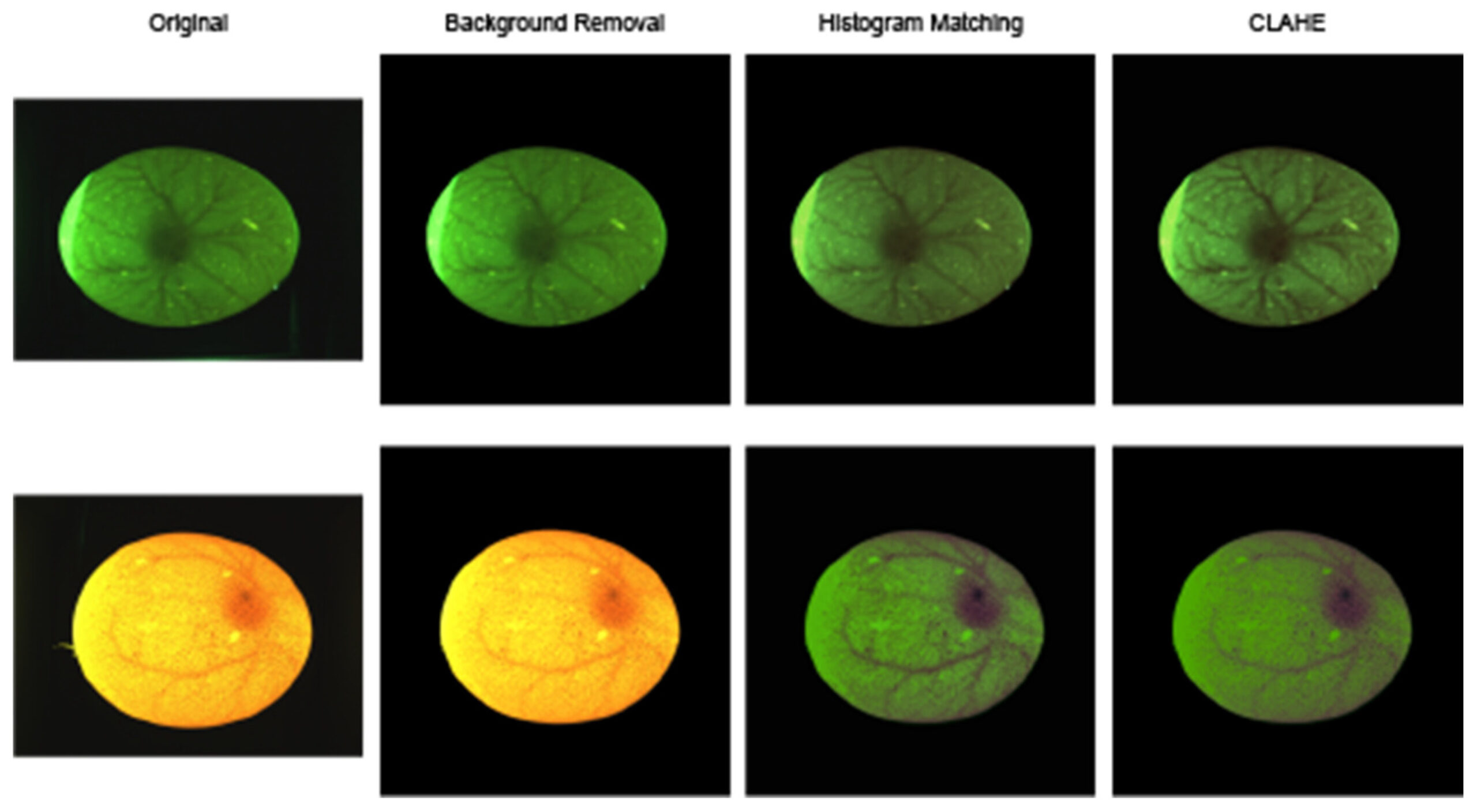
Рисунок 5. Конвейер предварительной обработки изображений. Показаны два репрезентативных изображения просвеченных яиц на разных этапах предварительной обработки, включая удаление фона, сопоставление гистограмм и повышение контрастности с использованием CLAHE. Различия в цвете между панелями отражают вариации светопропускания скорлупы и влияние этапов предварительной обработки. Они не представляют собой отдельные экспериментальные группы.
-
Изменение размера и добавление отступов;
-
Удаление фона;
-
Сопоставление гистограмм;
-
Снижение шума и адаптивное выравнивание гистограммы с ограничением контраста (CLAHE) [15].
Первым шагом было изменение размера и добавление отступов к изображениям в соответствии с входными размерами предварительно обученных моделей, описанных далее в этом исследовании. Затем было выполнено удаление фона с помощью модели сегментации, специально обученной для этой задачи.
Для учета различий во внешнем виде яичной скорлупы было применено сопоставление гистограмм. Некоторые яйца имели оранжевый оттенок, в то время как другие выглядели темно-зелеными; кроме того, некоторые изображения были слегка переэкспонированы, а другие — недоэкспонированы. Для устранения этих различий было использовано цветовое пространство CIELAB, которое разделяет изображение на компоненты L* (яркость), a* (зеленый-красный) и b* (синий-желтый). Затем сопоставление гистограмм было использовано для стандартизации цвета изображения и условий освещения.
На многих яйцах наблюдался значительный шум в виде небольших темных пятен или участков, где свет неравномерно проникал в скорлупу. Для уменьшения этого эффекта применялись морфологические операции открытия и закрытия, за которыми следовал медианный фильтр размытия. После уменьшения шума применялся CLAHE в меньшем масштабе для улучшения видимости сосудов при одновременном ограничении усиления остаточного шума [15]. Из-за высокого уровня шума на некоторых изображениях CLAHE нельзя было применять напрямую, так как в противном случае он усиливал бы шум в той же степени, что и значимые признаки.
Поскольку некоторые части набора данных содержали сильный шум в виде светлых и темных пятен, которые заслоняли эмбрион, был выполнен этап очистки (рис. 6). Изображения в верхнем ряду рисунка иллюстрируют примеры, которые были исключены, а нижний ряд показывает репрезентативные изображения, которые были сохранены в очищенном наборе данных.
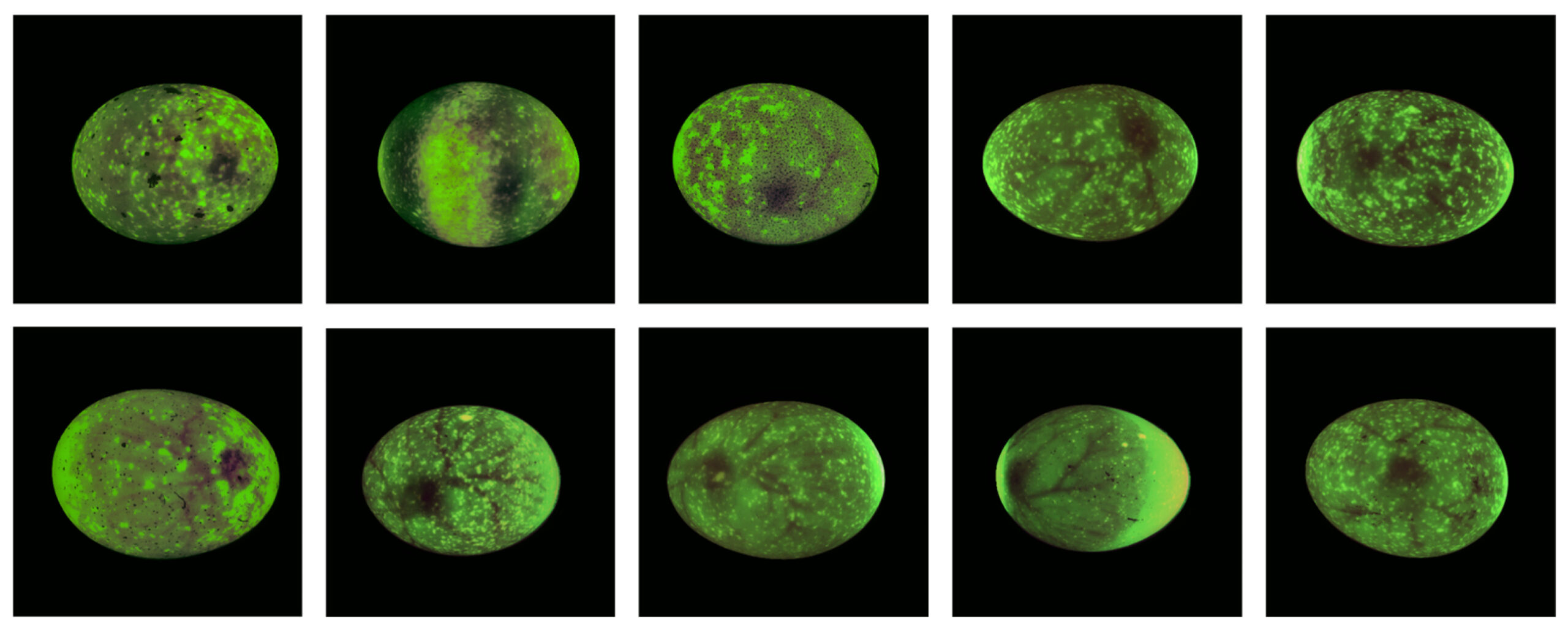
Рисунок 6. Примеры очистки изображений. В верхнем ряду показаны изображения, исключенные после ручной визуальной проверки из-за сильного шума или нечеткого отображения эмбрионов, а в нижнем ряду — репрезентативные изображения с 5-го дня, которые были сохранены в анализируемом наборе данных.
Как показано в таблице 2 , в общей сложности 21 серия изображений (соответствующая 72 изображениям) была удалена, поскольку эмбрионы были скрыты темными или светлыми пятнами. Эта фильтрация проводилась путем ручной визуальной проверки. В анализ машинного обучения были включены только яйца с полными сериями изображений. Хотя некоторые изображения в наборе данных все еще содержали шум, несколько были достаточно четкими для использования, как показано на рисунке 6. Примечательно, что данные, собранные для породы Вельзумер, показали особенно высокий уровень шума.
Таблица 2. Количество серий изображений (яиц) с подтвержденными метками самца и самки, включенных в анализ машинного обучения после удаления 21 серии (72 изображения) путем ручной визуальной проверки. Процент удаленных серий показан для каждой породы и пола.
| Порода | Мужской | Процент удаленных | Женский | Процент удаленных |
|---|---|---|---|---|
| Велсумер | 31 | 13,9% | 16 | 30,4% |
| Кремовый Легбар | 59 | 15,7% | 52 | 20,0% |
| Скрещивание | 16 | 11,1% | 14 | 12,5% |
| Сумма | 106 | 14,5% | 82 | 21,2% |
2.4. Модели машинного обучения
Модели машинного обучения были реализованы в TensorFlow (версия 2.13.0) [ 16 ] с использованием API Keras [17]. Предварительная обработка изображений проводилась с помощью OpenCV (версия 4.9.0) [18] и Scikit-image (версия 0.23.2) [19].
Для анализа как пространственных, так и временных характеристик последовательностей изображений использовалась комбинация моделей на основе сверточной нейронной сети (CNN) и рекуррентной нейронной сети (RNN). Учитывая относительно небольшой набор данных, были выбраны предварительно обученные модели с меньшим количеством параметров, чтобы снизить риск переобучения и использовать перенос обучения с больших наборов данных изображений. Модели на основе CNN были инициализированы предварительно обученными на ImageNet [20] и впоследствии дообучены для данного набора данных.
Оценивались модели ResNet50V2 [21], InceptionV3 [22] и DenseNet121 [23], содержащие приблизительно 25,6, 23,9 и 8,1 миллиона параметров соответственно, как показано на рисунке 7. Для оценки влияния выбора модели на производительность оценивались несколько архитектур CNN, а не для их объединения в одну модель. Несколько архитектур CNN использовались для оценки производительности моделей в различных базовых архитектурах, а не для их объединения в одну прогностическую модель.
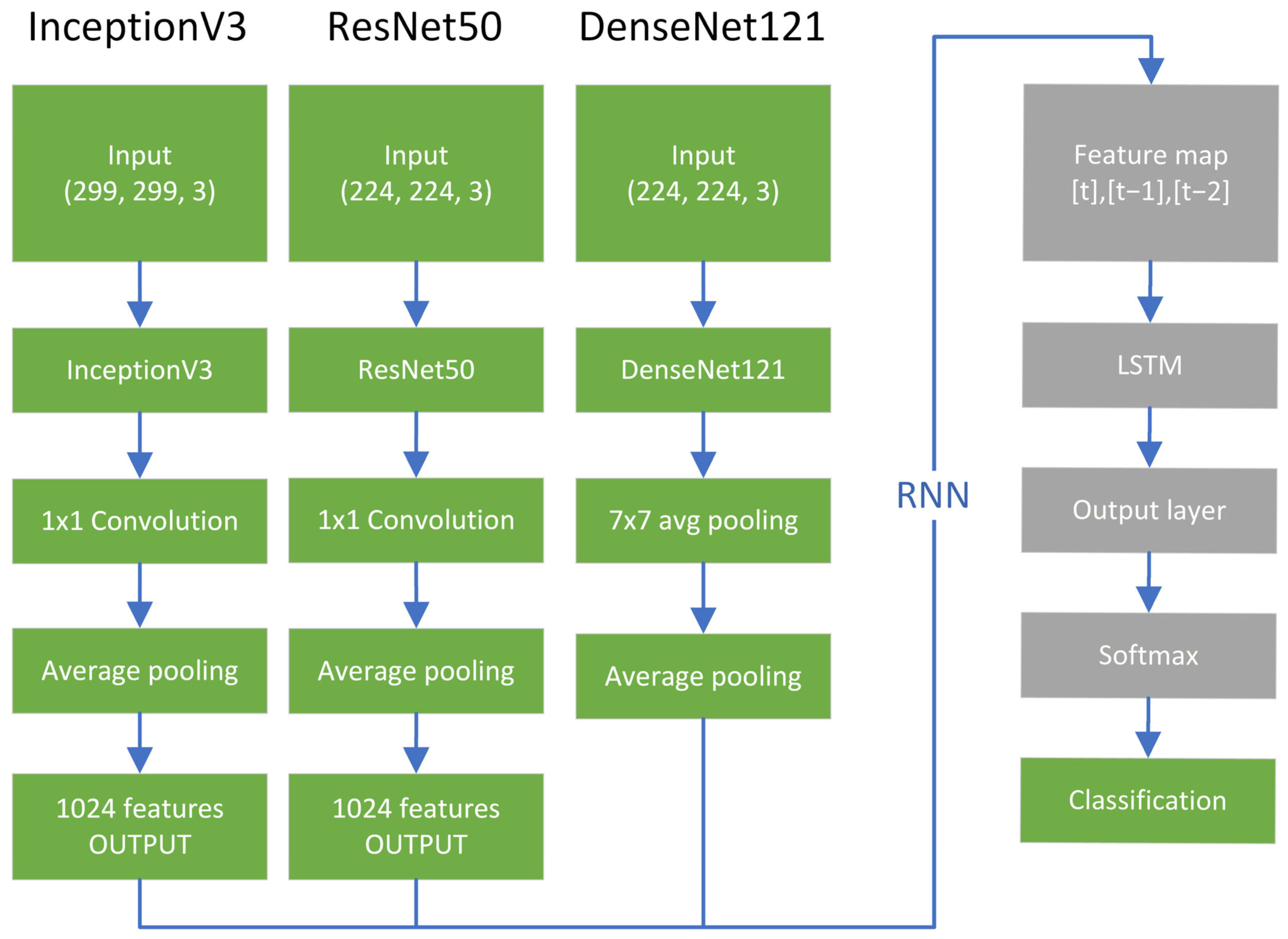
Рисунок 7. Архитектура гибридной модели CNN–RNN, иллюстрирующая сочетание извлечения пространственных признаков и моделирования временной последовательности. Модели оценивались независимо и не были объединены в единую архитектуру.
В процессе обучения применялось расширение данных для повышения устойчивости модели. Конвейер расширения данных включал случайные горизонтальные и вертикальные отражения, повороты, масштабирование и смещения, что вносило вариативность в ориентацию и масштаб изображений. Это было особенно актуально, учитывая вариативность ориентации яиц.
Яйца разных пород различались по цвету скорлупы и характеристикам поверхности. В частности, определить пол у кур породы Вельзумер было сложнее по оперению. Для решения этой проблемы были созданы два набора данных: один, включающий все три породы, и другой, исключающий яйца Вельзумер. Все три базовые модели были оценены на обоих наборах данных.
После оценки базовых сверточных нейронных сетей (CNN) каждая модель была расширена за счет рекуррентной нейронной сети (RNN) в каскадной архитектуре, аналогичной ранее предложенным структурам CNN–RNN. Такой подход был мотивирован более ранними исследованиями, которые подчеркнули временной аспект эмбрионального развития как перспективное направление для будущих исследований. Итоговая карта признаков, полученная из базовых CNN, была обработана с использованием слоя GlobalAveragePooling2D. Для InceptionV3 и ResNet50 был применен дополнительный слой Conv2D размером 1 × 1 для обеспечения размерной согласованности.
В этой модели сверточная нейронная сеть (CNN) извлекла карты признаков, состоящие из 1024 признаков, которые затем были переданы рекуррентной нейронной сети (RNN) в виде последовательности из трех карт признаков, что в общей сложности дало 3072 признака.
Для оценки эффективности различных базовых моделей в сочетании с RNN к базовым моделям CNN были применены идентичные параметры, в то время как гиперпараметры RNN были настроены эмпирически в ходе предварительных экспериментов. Набор данных был разделен на 70% для обучения, 15% для валидации и 15% для тестирования. Разделение проводилось на уровне яиц, чтобы гарантировать, что изображения одного и того же яйца не будут присутствовать в нескольких подмножествах.
Эффективность модели оценивалась с помощью точности, анализа матрицы ошибок, F1-меры и коэффициента корреляции Мэтьюса (MCC).
3. Результаты
3.1. Базовая модель CNN
В целом, точность базовых моделей CNN для полного набора данных лишь немного превышала случайный уровень, в то время как производительность на сокращенном наборе данных (исключая яйца Welsumer) была еще ниже. Среди базовых моделей InceptionV3, обученная на полном наборе данных за 6 дней, достигла наивысшей средней точности, составив 64,3%. Эта модель также получила показатель F1, равный 67,9%, что указывает на сбалансированную производительность между точностью и полнотой. Лучшим отдельным запуском среди всех вариантов базовых CNN также стала модель InceptionV3, обученная на полном наборе данных за 6 дней, которая достигла точности 71,4% на тестовом наборе (рисунок 8).
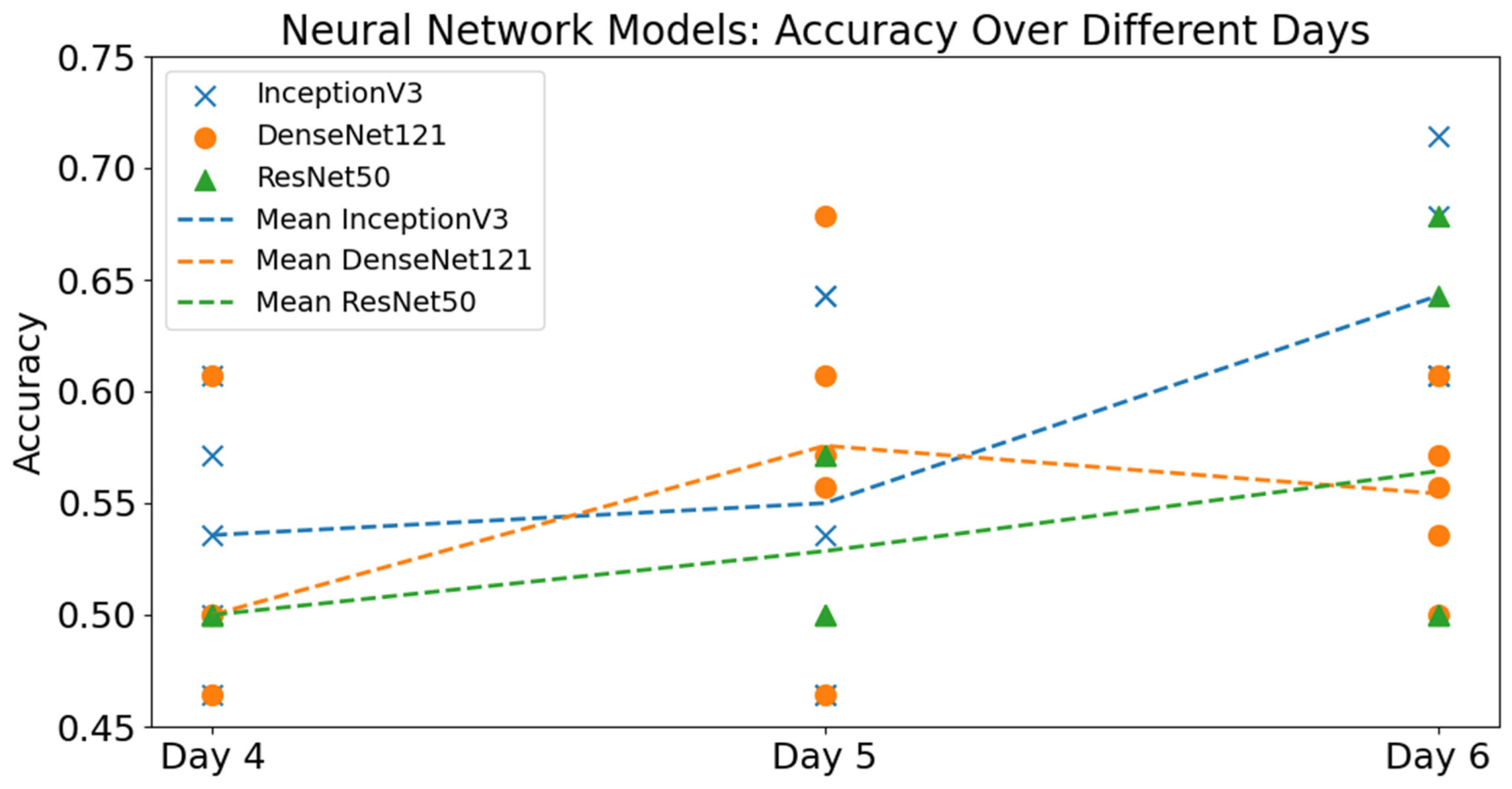
Рисунок 8. Сводные результаты по всем трем моделям CNN (ResNet50V2, InceptionV3, DenseNet121) и всем дням инкубации для полного набора данных, включающего все три породы.
3.2. Гибридная модель CNN-RNN
Средняя точность моделей CNN-RNN не показала существенного улучшения по сравнению с базовыми моделями CNN. Аналогично результатам CNN, InceptionV3 и DenseNet121 продемонстрировали более высокую производительность на полном наборе данных, в то время как ResNet50 показал лучшие результаты на подмножестве, исключающем яйца Welsumer. В целом, лучшей моделью оказалась DenseNet121, хотя модели на основе InceptionV3 показали более стабильную производительность в разных запусках, что привело к более высокой средней точности.
Производительность гибридных моделей CNN–RNN значительно варьировалась. На полном наборе данных ResNet50 достигла лишь 50% точности, в основном потому, что классифицировала почти все яйца как мужские. Это также подтверждается значением MCC, близким к нулю, что свидетельствует об отсутствии значимой прогностической способности. В отличие от этого, на сокращенном наборе данных ResNet50 показала более высокую точность, в то время как DenseNet121 продемонстрировала тенденцию к чрезмерной классификации яиц как мужских (Рисунок 9).
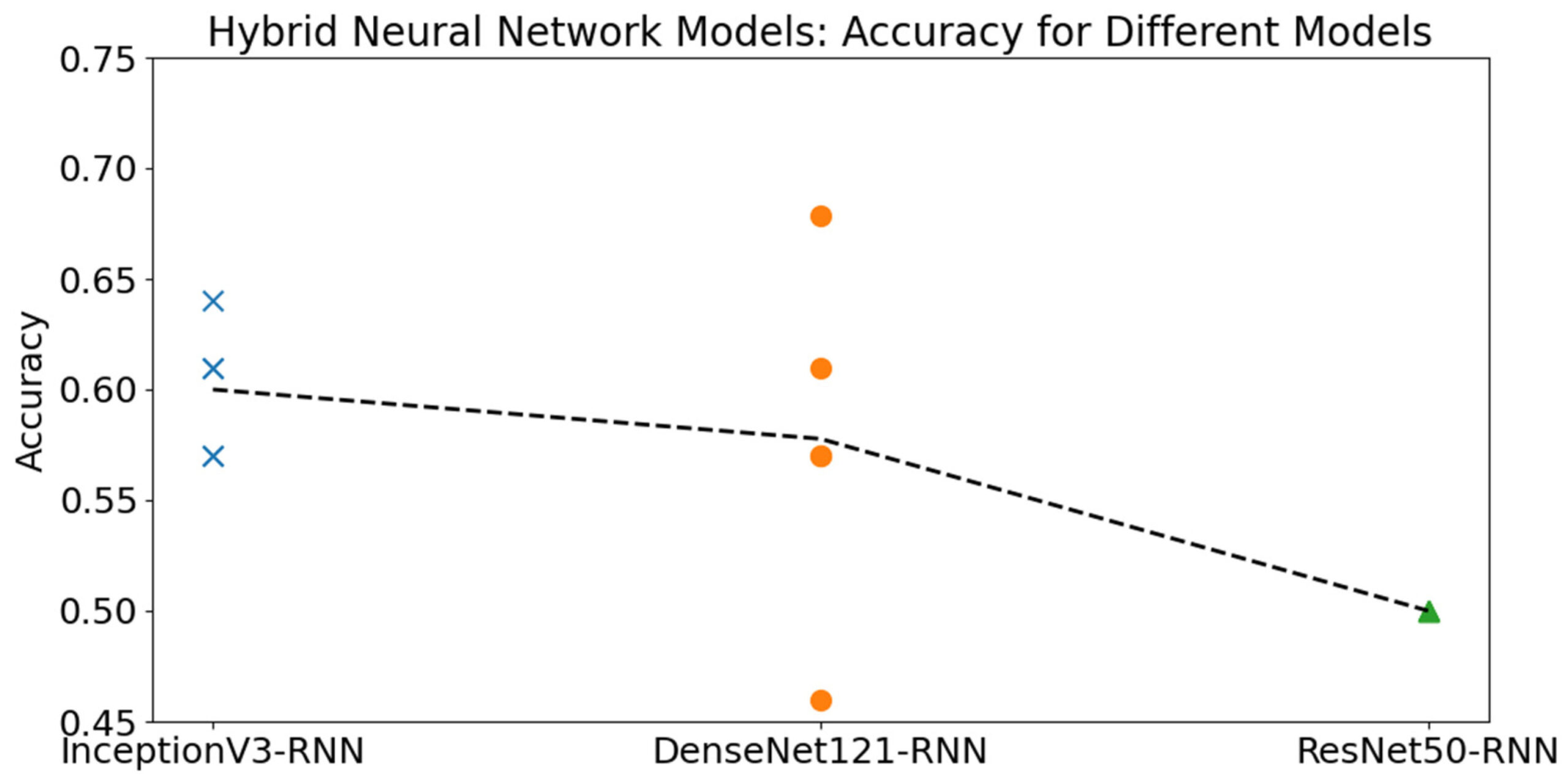
Рисунок 9. Точность трех гибридных моделей CNN–RNN в пяти независимых запусках. Символы представляют отдельные запуски для каждой архитектуры модели: кресты обозначают InceptionV3-RNN, точки — DenseNet121-RNN, а треугольник — ResNet50-RNN. Пунктирная линия соединяет средние значения точности для каждой модели.
Для более детальной оценки производительности модели на рисунке 10 показана матрица ошибок для модели с наилучшими показателями . На основе этой матрицы коэффициент корреляции Мэтьюса (MCC) составил приблизительно 0,31, что указывает на умеренную, но неслучайную прогностическую эффективность. Также можно наблюдать некоторый дисбаланс в прогнозах классов.
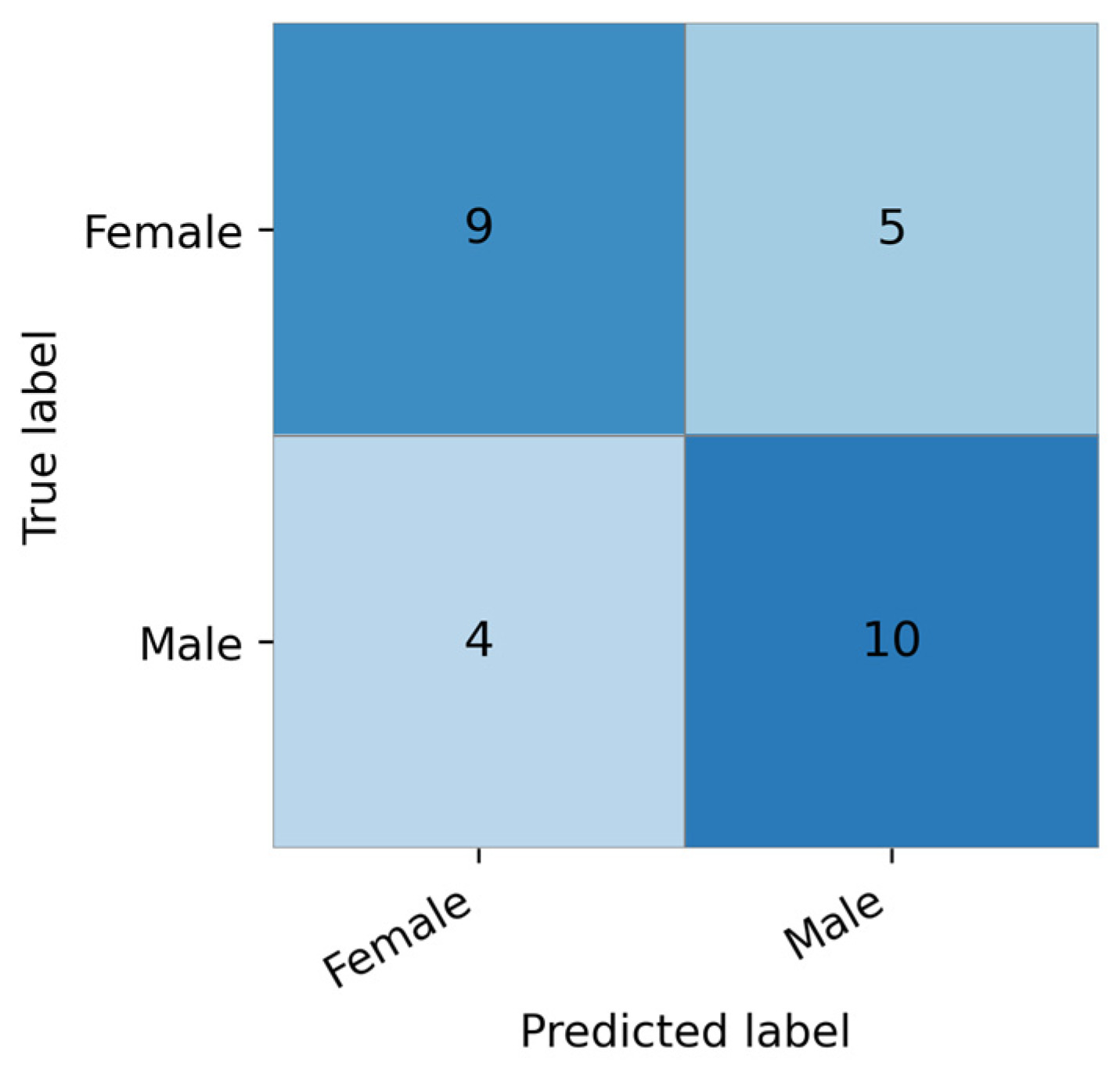
Рисунок 10. Матрица ошибок для модели с наилучшими показателями, оцененная на тестовом наборе данных.
4. Обсуждение
В данном исследовании изучалась возможность классификации пола куриных яиц на ранних стадиях развития с использованием сверточных нейронных сетей (CNN) и гибридной архитектуры CNN-RNN. Хотя несколько моделей достигли точности около 60%, положительное значение MCC (≈0,31) дополнительно подтверждает, что наиболее эффективная модель улавливает значимые закономерности в данных, хотя общая производительность остается скромной по сравнению с предыдущими исследованиями. Этот результат требует более глубокого изучения набора данных, методологии и последствий. Результаты следует интерпретировать как предварительные и отражающие сложность задачи в реалистичных условиях.
Наилучшие результаты показала модель InceptionV3 CNN, обученная на изображениях 6-го дня, достигнув точности 71,4% и показателя F1 67,9%. Однако включение временной информации с помощью RNN не улучшило производительность — фактически, модели CNN-RNN в среднем показали худшие результаты. Это говорит о том, что для ранних стадий визуализации (4–6 дни) статические пространственные характеристики могут быть более информативными, чем временная динамика.
Ограниченный размер и качество набора данных, включая шум от естественной изменчивости яиц и непостоянные условия освещения, вероятно, ограничили производительность модели. Хотя во время получения изображений использовался контролируемый источник света с фиксированной длиной волны, вариации цвета и структуры скорлупы влияют на то, как свет проходит через яйцо. Более темные или более неоднородные скорлупы поглощают и рассеивают больше света, снижая видимость эмбриональных структур и увеличивая изменчивость изображения.
Этот эффект был особенно заметен для породы Вельзумер, у которой наблюдался более высокий уровень шума из-за более темного и изменчивого внешнего вида скорлупы. С учетом этих ограничений результаты следует интерпретировать как предварительные, отражающие осуществимость подхода в реалистичных и неоднородных условиях, а не оптимизированную производительность. Сбор более крупных и сбалансированных наборов данных и проведение внешней валидации остаются важными направлениями для будущих исследований.
Несмотря на включение нескольких пород, удаление породы Вельзумер из набора данных не оказало существенного влияния на точность, что позволяет предположить, что задача классификации может быть применима к различным породам. Однако выводы остаются предварительными из-за относительно небольшого размера выборки и ограниченного числа полных серий изображений.
Более детальный анализ потенциальных различий, связанных с породами, например, с использованием анализа главных компонентов (PCA), был невозможен из-за ограниченного размера выборки для каждой породы и оставлен для будущих исследований. Набор данных несколько несбалансирован, в нем больше образцов самцов, чем самок, что может повлиять на производительность модели. Тем не менее, матрица ошибок указывает на относительно сбалансированную производительность классификации между двумя классами для модели с наилучшими показателями.
По сравнению с существующими исследованиями определения пола in ovo [2, 5, 6, 10, 24], данное исследование представляет уникальный набор данных, расширяющий рамки предыдущих исследований. Большая часть предыдущих работ была сосредоточена на коммерческих породах и однодневной визуализации; напротив, наш набор данных включает в себя традиционные и помесные породы кур — в частности, Welsumer, Cream Legbar и помесь Cream Legbar/Smålandshöna — что добавляет ценную генетическую и яичную изменчивость.
Кроме того, изображения были собраны в течение трех последовательных дней (дни 4, 5 и 6), что позволило провести временной анализ с помощью архитектур CNN-RNN. Хотя это временное моделирование не улучшило производительность по сравнению с базовыми CNN, структура набора данных предоставляет редкую возможность изучать эмбриональное развитие во времени.
Система визуализации, основанная на камере Raspberry Pi и массиве зеленых светодиодов, была недорогой и легко воспроизводимой, что делало этот подход осуществимым для децентрализованных или небольших инкубаторов. Важно отметить, что данные собирались в условиях, близких к коммерческим, а пол цыплят определялся после вылупления по характеристикам оперения, что обеспечивало как соблюдение этических норм, так и точность маркировки. Эти особенности делают набор данных полезным ресурсом для будущих исследований, направленных на обобщение методов классификации пола для разных пород, условий визуализации и временных масштабов.
Наши результаты сопоставимы с результатами недавнего турецкого исследования, в котором использовались модели ResNet и Inception на наборе данных аналогичного размера, достигнув точности 67,6% с ResNet и 75,5% с Inception [6]. В отличие от этого, наша гибридная модель CNN-RNN достигла наилучшей точности в 66,7%, со средним значением 62,7% на подмножестве, исключенном по породам. Эти различия могут быть обусловлены вариациями в предварительной обработке, четкости изображения и настройке сети. В других исследованиях, сообщающих о более высокой точности (до 89,0%), часто использовались более крупные, более контролируемые наборы данных или продвинутая обработка изображений, такая как гиперспектральная визуализация или морфологический анализ кровеносных сосудов [5, 10, 14, 24, 25, 26, 27].
Ранний, неинвазивный характер этого подхода остается многообещающим, несмотря на его нынешние ограничения. Возможность классифицировать пол уже на 4-й день, не вскрывая яйцо и не влияя на его жизнеспособность, хорошо согласуется с новыми правилами ЕС, запрещающими выбраковку цыплят [1, 2]. Хотя точность должна улучшиться, чтобы соответствовать коммерческим стандартам, наши результаты подтверждают жизнеспособность машинного зрения как гуманной альтернативы, заслуживающей дальнейшего развития.
Предыдущие исследования предполагают, что развитие кровеносных сосудов у эмбрионов может различаться между полами из-за вариаций метаболической активности и динамики роста на ранних стадиях инкубации. Эти различия могут проявляться в виде вариаций плотности сосудов, характера ветвления и пространственной организации, которые можно зафиксировать с помощью анализа изображений. Хотя точные биологические механизмы остаются недостаточно изученными, это дает правдоподобную основу для использования сосудистых паттернов в качестве входных признаков для классификации пола [5].
Дальнейшие исследования должны быть сосредоточены на улучшении качества изображений и размера набора данных. Это может включать автоматизированные системы получения изображений для обеспечения равномерного освещения, а также мультиспектральную или гиперспектральную съемку для повышения контраста сосудов. Кроме того, изучение алгоритмов обнаружения объектов в реальном времени, таких как варианты YOLOv7 [28], может обеспечить практическое применение в условиях рыбоводных хозяйств. Наконец, более узкие интервалы выборки между днями съемки могут позволить более эффективно использовать временные данные, потенциально повышая производительность на основе RNN.
В заключение, хотя временной аспект не привел к улучшению производительности в данном исследовании, базовая модель CNN показала обнадеживающие результаты. При наличии более чистого и крупного набора данных, а также при дальнейшем совершенствовании архитектуры модели, неинвазивное определение пола на ранних стадиях с помощью машинного зрения остается перспективным направлением исследований.
5. Выводы
В данном исследовании изучалось, может ли включение временной информации — посредством многодневных серий изображений и гибридной архитектуры CNN-RNN — улучшить неинвазивное определение пола на ранних стадиях развития куриных яиц. Несмотря на новизну этого подхода, результаты показали, что базовая модель CNN превзошла гибридную модель, достигнув точности до 71,43% по сравнению с 67,85% для CNN-RNN. Скромные результаты, вероятно, обусловлены небольшим размером набора данных и значительным шумом в изображениях.
Отличительной особенностью этой работы является использование традиционных и помесных линий кур, а не промышленных бройлеров, а также сбор данных изображений в течение трех дней инкубации. Такой подход вносит естественную изменчивость и более точно отражает условия, выходящие за рамки контролируемых промышленных условий, предоставляя ценный пример для проверки возможности обобщения результатов.
Дальнейшая работа должна быть сосредоточена на создании более крупного и качественного набора данных с более равномерным освещением и меньшим количеством артефактов. Кроме того, более узкие интервалы между снимками и модели обнаружения объектов в реальном времени могут лучше использовать временные изменения в сосудистых рисунках. Хотя подход CNN-RNN не показал повышения производительности в этом исследовании, общие результаты подтверждают потенциал машинного зрения как неинвазивного, масштабируемого и этически обоснованного метода раннего определения пола цыплят. В будущих исследованиях также следует использовать методы объяснимости, такие как Grad-CAM или SHAP, чтобы лучше понять, какие характеристики изображения влияют на прогнозы модели.
Литература:
- Coppola, F.; Paci, G.; Profeti, M.; Mancini, S. Stop culling male layer-type chick: An overview of the alternatives and public perspective. World’s Poult. Sci. J. 2024, 80, 611–631. [Google Scholar] [CrossRef]
- Krautwald-Junghanns, M.-E.; Cramer, K.; Fischer, B.; Förster, A.; Galli, R.; Kremer, F.; Mapesa, E.U.; Meissner, S.; Preisinger, R.; Preusse, G.; et al. Current approaches to avoid the culling of day-old male chicks in the layer industry, with special reference to spectroscopic methods. Poult. Sci. 2018, 97, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Steiner, G.; Bartels, T.; Stelling, A.; Krautwald-Junghanns, M.-E.; Fuhrmann, H.; Sablinskas, V.; Koch, E. Gender determination of fertilized unincubated chicken eggs by infrared spectroscopic imaging. Anal. Bioanal. Chem. 2011, 400, 2775–2782. [Google Scholar] [CrossRef] [PubMed]
- Galli, R.; Preusse, G.; Uckermann, O.; Bartels, T.; Krautwald-Junghanns, M.-E.; Koch, E.; Steiner, G. In ovo sexing of domestic chicken eggs by Raman spectroscopy. Anal. Chem. 2016, 88, 8657–8663. [Google Scholar] [CrossRef] [PubMed]
- Jia, N.; Li, B.; Zhao, Y.; Fan, S.; Zhu, J.; Wang, H.; Zhao, W. Exploratory study of sex identification for chicken embryos based on blood vessel images and deep learning. Agriculture 2023, 13, 1480. [Google Scholar] [CrossRef]
- Asil, U.; Nasibov, E. Sex detection in the early stage of fertilized chicken eggs via image recognition. Int. J. Comput. Sci. Inf. Technol. 2023, 15, 19–26. [Google Scholar]
- Xie, C.; Wang, T.; Yang, C. A review of recent advances in in ovo sexing of chicken embryos using optical sensing techniques. Poult. Sci. 2023, 102, 102906. [Google Scholar] [CrossRef] [PubMed]
- Ekebo Fjäderfä. Available online: https://www.ekebofjaderfa.com/ (accessed on 26 June 2024).
- Brinsea Products Ltd. Ova-Easy 190 Advance Series II Incubator; Brinsea Products Ltd.: Weston-Super-Mare, UK, 2024. [Google Scholar]
- Zhu, Z.; Ye, Z.F.; Tang, Y. Nondestructive identification for gender of chicken eggs based on GA-BPNN with double hidden layers. J. Appl. Poult. Res. 2021, 30, 100203. [Google Scholar] [CrossRef]
- Archer, G.S.; Mench, J.A. Natural incubation patterns and the effects of exposing eggs to light at various times during incubation on post-hatch fear and stress responses in broiler chickens. Appl. Anim. Behav. Sci. 2014, 152, 44–51. [Google Scholar] [CrossRef]
- Rozenboim, I.; Piestun, Y.; Mobarkey, N.; Barak, M.; Hoyzman, A.; Halevy, O. Monochromatic light stimuli during embryogenesis enhance embryo development and posthatch growth. Poult. Sci. 2004, 83, 1413–1419. [Google Scholar] [CrossRef] [PubMed]
- Sobolewska, A.; Elminowska-Wenda, G.; Bogucka, J.; Szpinda, M.; Walasik, K.; Dankowiakowska, A.; Jóźwicki, W.; Wiśniewska, H.; Bednarczyk, M. The effect of two different green lighting schedules during embryogenesis on myogenesis in broiler chickens. Eur. Poult. Sci. 2019, 83, 1–14. [Google Scholar] [CrossRef]
- El-Sabrout, K.; Khalil, M.H. Effect of LED lighting on hatchability and chick performance of chicken eggs. Pak. J. Zool. 2017, 49, 2323–2325. [Google Scholar] [CrossRef]
- Zuiderveld, K. Contrast limited adaptive histogram equalization. In Graphics Gems IV; Heckbert, P.S., Ed.; Academic Press: San Diego, CA, USA, 1994; pp. 474–485. [Google Scholar]
- TensorFlow. Available online: https://www.tensorflow.org/ (accessed on 27 June 2024).
- Chollet, F. Keras: The Python Deep Learning API. Available online: https://keras.io (accessed on 27 June 2024).
- OpenCV. Open Source Computer Vision Library. Available online: https://opencv.org (accessed on 5 September 2025).
- van der Walt, S.; Schönberger, J.L.; Nunez-Iglesias, J.; Boulogne, F.; Warner, J.D.; Yager, N.; Gouillart, E.; Yu, T. Scikit-Image: Image Processing in Python. Available online: https://scikit-image.org (accessed on 5 September 2025).
- Deng, J.; Dong, W.; Socher, R.; Li, L.-J.; Li, K.; Li, F.-F. ImageNet: A large-scale hierarchical image database. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Miami, FL, USA, 20–25 June 2009. [Google Scholar]
- He, K.; Zhang, X.; Ren, S.; Sun, J. Identity mappings in deep residual networks. In Proceedings of the European Conference on Computer Vision (ECCV), Amsterdam, The Netherlands, 11–14 October 2016. [Google Scholar]
- Szegedy, C.; Vanhoucke, V.; Ioffe, S.; Shlens, J.; Wojna, Z. Rethinking the Inception architecture for computer vision. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Las Vegas, NV, USA, 27–30 June 2016. [Google Scholar]
- Huang, G.; Liu, Z.; van der Maaten, L.; Weinberger, K.Q. Densely connected convolutional networks. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Honolulu, HI, USA, 21–26 July 2017. [Google Scholar]
- Tang, Y.; Hu, Q.; Wang, Q.; Zhu, Z. Sex identification of chicken eggs based on blood line texture features and GA-BP neural network. J. Huazhong Agric. Univ. 2018, 37, 130–135. [Google Scholar]
- Weissmann, A.; Reitemeier, S.; Hahn, A.; Gottschalk, J.; Einspanier, A. Sexing domestic chicken before hatch: A new method for in ovo gender identification. Theriogenology 2013, 80, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Göhler, D.; Fischer, B.; Meissner, S. In-ovo sexing of 14-day-old chicken embryos by pattern analysis in hyperspectral images. Poult. Sci. 2017, 96, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Alin, K.; Fujitani, S.; Kashimori, A.; Suzuki, T.; Ogawa, Y.; Kondo, N. Non-invasive broiler chick embryo sexing based on opacity value of incubated eggs. Comput. Electron. Agric. 2019, 158, 30–35. [Google Scholar] [CrossRef]
- Wang, C.-Y.; Bochkovskiy, A.; Liao, H.-Y.M. YOLOv7: Trainable Bag-of-Freebies Sets New State-of-the-Art for Real-Time Object Detectors. Available online: ссылка (accessed on 14 September 2025).
Авторы: Joel Andersson, Per Magnusson, Fredrik Frisk


